This is the next-to-last full contribution of photos I have, so please send in good wildlife photos if you got ’em. Thanks!
Today we have a text-and-photo discussion from reader Athayde Tonhasca Júnior. Athayde’s text is indented, and you can enlarge the photos by clicking on them.
Complex marriage arrangements
In his Recollections of the development of my mind and character, autobiographical reflections edited by his son Francis in 1887, Charles Darwin remarked: “I do not think anything in my scientific life has given me so much satisfaction as making out the meaning of the structure of heterostylous flowers.” If Darwin chose explaining heterostyly as one of his greatest scientific achievements among all his pivotal contributions, perhaps we could share his sense of gratification—once we know what heterostyly is.
Flowering plants are predominantly hermaphroditic: their flowers produce female (pistils) and male (stamens) sexual organs. The pistil comprises the ovary, the style (a pillar-like stalk through which pollen germinates to reach the ovary), and the stigma (a sticky tip at the top of the style that receives pollen). The stamen has a filament that supports the anthers, where pollen is produced.
Parts of a hermaphroditic flower © Anjubaba, Wikimedia Commons:
{kind=link}

From the Greek heteros (different) and stylos (pillar), heterostyly is the condition where flowers of a species fall into one of two types, or morphs (sometimes three, but these cases are not discussed here): one where the female reproductive parts are high up in the flower and the male organs are in the bottom, and the other with the opposite arrangement. Heterostyly is a particular type of herkogamy, which is the spatial separation of reproductive structures.
Primroses, cowslips and oxlips (Primula spp.) are the best known examples of this disposition of sex organs. Their flowers come in two morphs, each in different plants: ‘pins’, which have elongated styles and short stamens, and ‘thrums’, with short styles and elongated stamens. ‘Pins’ and ‘thrums’ are also known as L-morphs (long styles) and S-morphs (short styles), respectively.
Primrose morphs: (L), a pin flower, named after ‘the similarity of the stigma to the head of a pin’ (Darwin, 1862) © Velela; (R): a thrum flower (in weaving, ‘thrum’ is a fringe of threads) © Velela, Wikimedia Commons:
{kind=link}
{kind=link}

Primroses came to Darwin’s attention through his friend and mentor John S. Henslow, the botanist who had helped his pupil secure the post of naturalist aboard HMS Beagle. The ‘pins’ and ‘thrums’ phenomenon, including its terminology, had been known for more than a century before Darwin (Gilmartin, 2015), but nobody could explain the reasons for such apparently odd assembly.
As styles and stamens of primroses and other heterostylous plants are at different heights in each flower, their stigmas and anthers (the bits that receive and produce pollen) are also out of kilter. But here’s the important thing: stigmas in one morph are at about the same height of the anthers on the reciprocal morph. Darwin suspected that these dispositions were adaptations to cross-fertilisation.
Flowers of the common primrose (P. vulgaris). Pin or pin-eyed, pinheaded (L), and thrum or thrum-eyed. 1: anthers, 2: stigma. © Enrico Blasutto, Wikimedia Commons:
{kind=link}
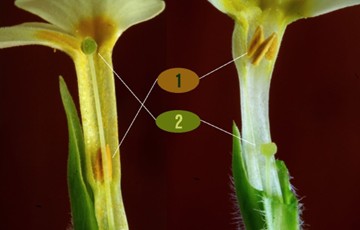
In his typically fastidious fashion, and barely one year after the publication of On The Origin of Species, Darwin found himself busy in his garden experimenting with cowslips (P. veris) to test his hypothesis. Darwin covered some cowslips with netting to prevent insects from reaching the flowers. He then hand-pollinated them in the following combinations: thrum flowers pollinated with pin pollen and vice versa, thrum flowers pollinated with thrum pollen, and pin flowers pollinated with pin pollen. When pollinating the same morphs, Darwin used pollen from different plants to avoid possible effects of inbreeding. After plants matured, he counted and weighed the seeds as measures of reproductive success. The results: inter-morph crosses, i.e., between pin and thrum flowers, produced more and heavier seeds; Darwin termed these cases ‘legitimate unions’. Crosses within morphs, be it in the same plant or between plants, were not as successful (‘illegitimate unions’). The benefit which heterostyled dimorphic plants derive from the existence of the two forms is sufficiently obvious, namely, the intercrossing of distinct plants being thus ensured. Nothing can be better adapted for this end than the relative positions of the anthers and stigmas in the two forms (Darwin, 1877).
Pathways of P. vulgaris intra- and inter-floral pollen movement from pin or long-styled (L, upper panel) and thrum or short-styled (S, lower panel) pollen donors. Solid lines indicate transfer of compatible pollen between reciprocal organs; dashed and dotted lines indicate transfer of incompatible pollen within flowers and between non-reciprocal organs of the same morph, respectively © Keller et al., 2014:
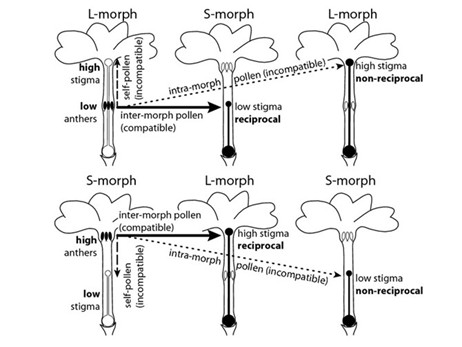
For Darwin (1864), Nature has ordained a most complex marriage arrangement for primroses, which he suspected had to do with insect pollination.
It had long been known that pollen grains usually stick to a specific body part of an insect probing a flower; when the visitor goes to another one, pollination is more likely to occur if the location of the stigma matches the location of the anthers in the first flower. Darwin concluded, reasonably, that insects must be responsible for the ‘legitimate unions’ of primroses and other heterostylous plants. Transfer of pollen between plants with reciprocal position of their sexual organs (i.e., the female parts in a thrum flower being at the same level as male parts from a pin flower) is much more likely to occur if carried out by flower-visiting insects. This process is helped by the tubular shape of most heterostylous flowers, which are usually visited by long-tongued pollinators such as bumble bees and moths that are able to probe deep in search of nectar.
Pollination of heterostylous flowers. Pollen collected from the short stamens in a long-styled morph (L) is deposited on the stigma of a short-styled morph (S); pollen from the S morph, stuck to different parts of the insect’s body, is transferred to the stigma of a L morph © Simón-Porcar et al., 2024:
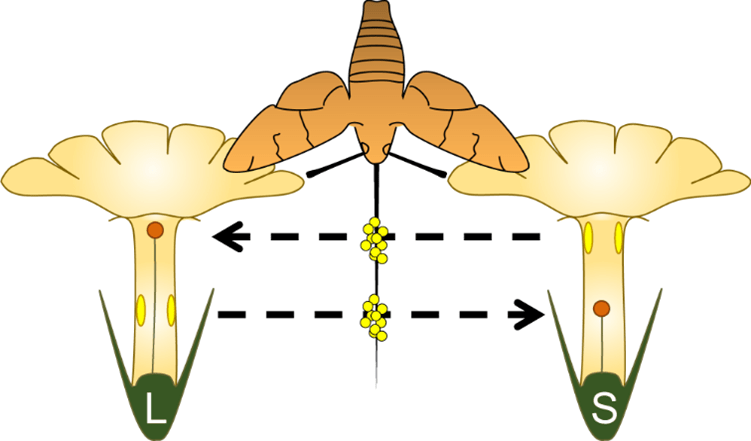
Subsequent work with several heterostylous species has supported Darwin’s cross-pollination hypothesis and added another quirk to the story: heterostyly is often associated with self-incompatibility, which are physiological processes that inhibit self-pollination. Heterostyly then not only promotes outcrossing, but also avoids inbreeding. This morphological arrangement is not that common but has been reported for at least 28 angiosperm families, an indication that it has evolved independently multiple times.
The humble cowslip, the main subject of Darwin’s scientific pursuits during the 1850s © BerndH, Wikimedia Commons:
{kind=link}

Despite lacking a genetic framework, not available until Gregor Mendel started playing with his peas, Darwin documented reduced fitness from selfing (self-fertilisation) for some 57 plant species he investigated at home (Barrett, 2010). He put much effort into examining plants’ reproductive biology and the form of their flowers to understand how they avoided the traps of selfing. His studies paid off. Dawin concluded that most hermaphroditic plants have developed barriers to avoid or reduce the possibility of self-fertilisation such as heterostyly and dichogamy, the temporal separation of male and female stages.
It’s understandable then that Darwin was so thrilled by cracking the function of heterostyly. He had added one more building block to his and Alfred Russel Wallace’s theory of evolution by natural selection, which remains the bedrock for the workings of the natural world.
Darwin’s greenhouse and garden where he carried countless experiments © Mario Modesto Mata, Wikimedia Commons:
{kind=link}

Wonderful post. Thanks, Athayde!
The wild English primrose is probably my favourite flower. I was born on the escarpment edging the north side of the Marlborough Downs, and in the deep coombes the chalk streams would flow down to the plain below. Always primroses to be found at their sides.
Very interesting discussion! Thanks!
Thanks so much for this information. Very informative.
“stigmas in one morph are at about the same height of the anthers on the reciprocal morph.”
Huh – I never knew that.
A take on popular slogans comes to mind from reading this :
Be like Darwin…
Or perhaps better and more edgy,..
What Would Darwin Do?
I never fail to learn something new and interesting from your contributions, so thank you!
Darwin was a very good experimentalist. His ability to use experiment to disentangle possibilities was extraordinary.
Excellent post!
Once again, thanks for an enlightening post.